Lorsque des gènes sont transmis d'une génération à une autre via la méiose et la fécondation, on parle de transfert vertical de gène (des ascendants vers les descendants). D'autres échanges génétiques peuvent avoir lieu entre deux organismes sans fécondation. On parle cette fois de transfert horizontal de gènes, rencontré souvent chez les micro-organismes (mais pas que) et intervient aussi dans les cas d'endosymbioses.
Les transferts horizontaux de gènes
Le transfert horizontal de gènes est un échange de matériels génétiques entre deux individus, pas forcément de la même espèce, sans fécondation ni reproduction. Ces transferts horizontaux de gènes sont possibles car l’ADN est une molécule universelle dont la structure est identique dans l’ensemble du monde vivant. Chez les bactéries, les transferts horizontaux sont très fréquents mais il peut aussi arriver que le transfert d’ADN se fasse entre une bactérie et un organisme eucaryote ou par l’intermédiaire de virus.
Il existe plusieurs modalités de transferts de gènes horizontaux entre les bactéries (parmi lesquelles la transformation bactérienne, la conjugaison bactérienne et la transduction bactérienne). Mais au cours de l'histoire évolutive des êtres vivants, des transferts de ce type ont aussi eu lieu entre espèces n'appartenant pas à la même lignée (entre virus et mammifère ou bactérie et chordé par exemple).
La transformation bactérienne
La transformation bactérienne consiste en l’incorporation de fragments d’ADN libre dans le milieu extérieur dans le génome d’une bactérie receveuse. Le matériel génétique est fixé par les bactéries receveuses au niveau de leur surface puis absorbé par l’intermédiaire de protéines spécialisées.
Ce phénomène peut avoir lieu naturellement lorsqu’une bactérie meurt et que son contenu se retrouve dans l’environnement. Les fragments d’ADN ainsi libérés sont ensuite reconnus et captés par d’autres bactéries vivantes puis éventuellement intégrés dans leur génome.
La transformation bactérienne peut aussi être induite en laboratoire et être utilisée comme outil de manipulation génétique. Dans ce cas, on réalise un choc thermique (après avoir perméabilisé la membrane à l’aide d’une solution de Ca2+) ou une électroporation en faisant passer un courant électrique dans le milieu de culture afin, dans les deux cas, de permettre à l’ADN de traverser la membrane.

La conjugaison bactérienne
La conjugaison bactérienne est le transfert de matériel génétique par contact direct entre deux bactéries par l’intermédiaire d’un pont cytoplasmique. La conjugaison bactérienne débute par une bactérie donneuse (F+) qui étend son pilus sexuel (des pili sexuels) vers une bactérie receveuse (F-). Une fois le contact établi, les deux bactéries vont se rapprocher l’une de l’autre. Lorsqu'elles sont assez proches, un pont cytoplasmique va se former entre les deux bactéries et permettre l’échange de matériel génétique.
La bactérie donneuse F+ peut transférer une partie de son ADN chromosomique (après avoir été répliqué) ou alors un plasmide (un petit ADN circulaire distinct du chromosome bactérien) après avoir été lui aussi répliqué.

La transduction bactérienne
La transduction bactérienne est le transfert de matériel génétique entre deux bactéries par l’intermédiaire d’un bactériophage (virus qui infecte uniquement les bactéries). Dans ce cas, le virus est le vecteur du transfert.
Les virus ne peuvent se reproduire qu’en utilisant la machinerie cellulaire de l’hôte qu’ils infectent. Lorsque le virus infecte la bactérie, le génome du phage va se répliquer et l’ADN bactérien va être dégradé par les enzymes virales. De nouvelles particules virales vont se former dont certaines en possession de fragments de l’ADN bactérien. La cellule bactérienne est ensuite détruite et les phages libérés pourront aller infecter d’autres bactéries.
Le phage ne possédant plus que de l’ADN bactérien aura perdu de sa virulence et donc sa capacité à tuer les bactéries. Lorsqu’il infecte une autre bactérie, il injecte l’ADN de la précédente bactérie dans la nouvelle bactérie et par recombinaison ira s’incorporer dans le nouveau génome.

Transfert horizontal de gènes vers les eucaryotes
Un exemple de transfert horizontal de gènes impliquant des eucaryotes concerne la syncytine qui est une protéine impliquée dans la formation du placenta chez la grande majorité des mammifères. Cette protéine permet la fusion entre cellules qui aboutit à des cellules géantes à plusieurs noyaux caractéristiques du placenta.
Le gène codant pour la syncytine possède une séquence très proche d'un gène codant pour une protéine de l'enveloppe du rétrovirus MSRV. Un rétrovirus est un virus dont le matériel génétique est constitué d'ARN et qui utilise la machinerie cellulaire de la cellule hôte qu'il infecte pour incorporer son matériel génétique dans celui de l'hôte et se reproduire. Cette protéine de l'enveloppe permet au rétrovirus MSRV de fusionner avec la cellule hôte afin de faire pénétrer son matériel génétique.

On note donc une similitude à la fois dans la nature de la syncytine au niveau du placenta et de la protéine de l'enveloppe du rétrovirus et dans leurs modes d'action. L'hypothèse envisagée est que le gène codant pour la syncytine est un héritage laissé il y a plusieurs millions d'années par ce rétrovirus qui aurait infecté l'ancêtre commun des mammifères et aurait permis l'apparition des mammifères placentaires.
Les endosymbioses
Les cas d'endosymbiose dans l'histoire évolutive des êtres vivants sont nombreux et décisifs puisqu'ils ont été à l'origine des mitochondries et des plastes dans les cellules eucaryotes. C'est la théorie endosymbiotique.
La théorie endosymbiotique
Des bactéries aérobies auraient pénétré par invagination des cellules eucaryotes primitives puis dégénérées pour devenir les mitochondries actuelles. Ensuite, certaines de ces cellules nouvellement dotées de mitochondries auraient à leur tour, par invagination, incorporées en elles d'autres bactéries photosynthétiques (les cyanobactéries) pour donner naissance aux chloroplastes et à la lignée verte.
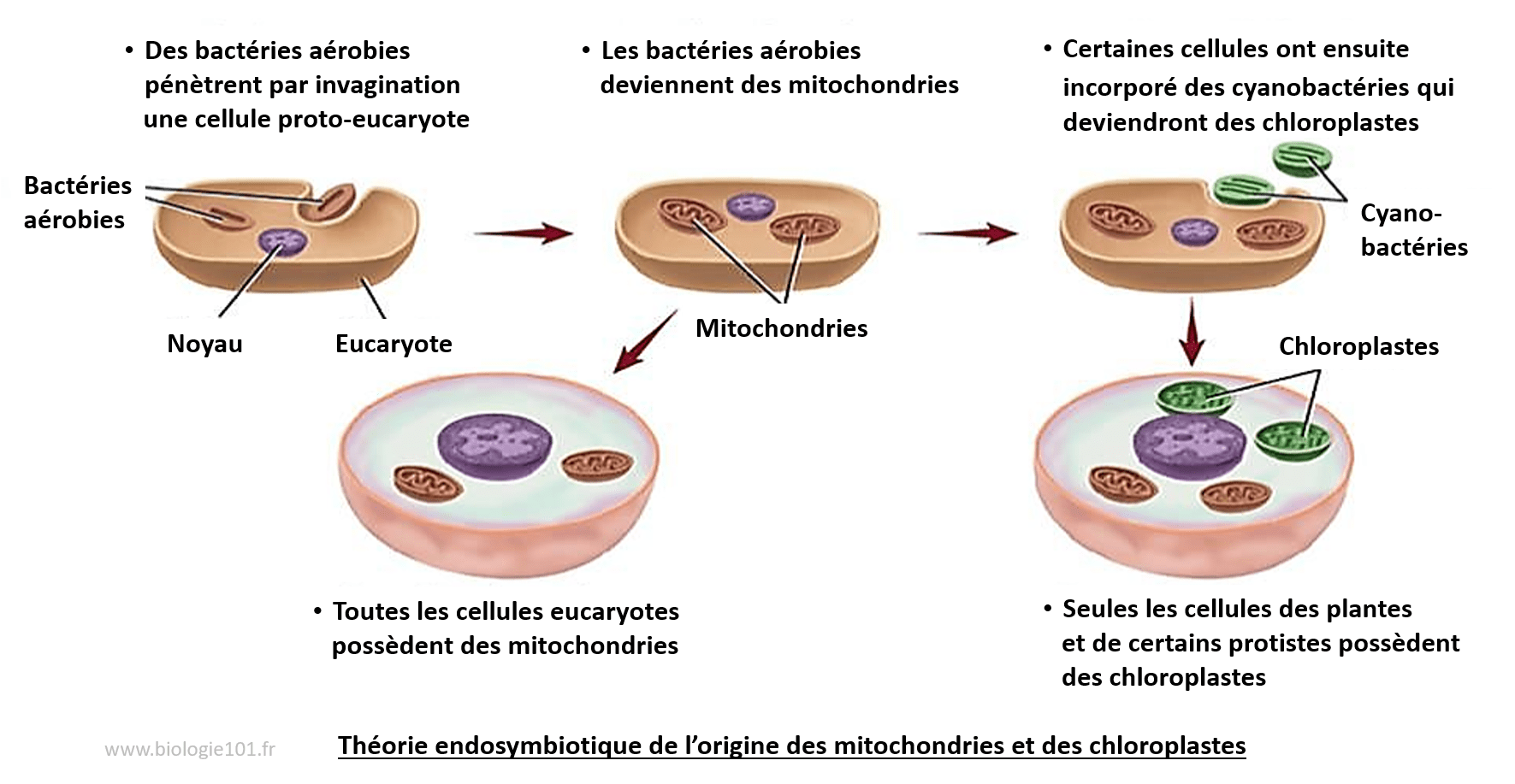
Les arguments pour une origine endosymbiotique des chloroplastes et des mitochondries sont les suivants :
- La taille des organites (1 à 5 μm) du même ordre de grandeur que celle d’une bactérie
- La présence de 2 membranes, une membrane interne dont la composition lipidique est proche de celle des procaryotes et une membrane externe semblable à la membrane unique de la cellule eucaryote qui a servi d’hôte
- Le processus de division des chloroplastes et des mitochondries est le même que celui des bactéries (la scissiparité)
- La présence d’ADN circulaire dans ces organites capable d’être transcrit et traduit de manière autonome
- Le génome de ces organites proche de celui des procaryotes
Transfert horizontal de gène entre le symbiote et l'hôte
Le génome des cellules eucaryotes possède de nombreux gènes proches de ceux des cellules procaryotes. L’explication est qu’au cours de l’évolution, il y a eu un transfert horizontal des gènes présents dans la cellule procaryote à l’origine des chloroplastes ou des mitochondries vers le noyau de la cellule eucaryote hôte.
De plus, plusieurs centaines de protéines sont traduites dans le noyau de la cellule et sont ensuite dirigées vers l’organite (le chloroplaste par exemple) afin d’assurer le fonctionnement de celui-ci, ce qui rend ces organites totalement dépendant de la cellule hôte.
![]()